Triploid Induction in Populus alba x P. glandulosa by Chromosome Doubling of Female Gametes
By Y. H. LI1), X. Y. KANG1),*), S. D. WANG1), Z. H. ZHANG1) and H. W. CHEN1)
Abstract
triploids. Triploids were obtained by chromosome dou-
Efficient methods for obtaining unreduced female
bling of female gametes from Morus alba L. (DWIVEDI et
gametes are needed for triploid induction in Populus
al., 1989) and P. tomentosa x P. bolleana (LI et al., 2001). alba x P. glandulosa. A cytological study of MMCs and
The incidence of triploid poplars, however, was low and
PMCs meiosis and staminate floral elongation revealed
consistent results were difficult to obtain in repeated
that there was a close association between MMCs meiot-
experiments. These studies identified the need to corre-
ic stage and male bud phenology during development
late meiotic stage of megaspore mother cells (MMCs)
under the same environment conditions. Female buds of
with colchicine treatment. Unfortunately, MMCs meio-
Populus alba x P. glandulosa were treated with 0.5 %
sis can only be observed through paraffin sectioning,
colchicine solution at the selected meiotic stages based
which is a slow and destructive process. This study was
on male bud phenology. The treated buds were pollinat-
conducted to examine the feasibility of an alternative,
ed with pollen of P. tomentosa. A high rate of unreducedfemale gametes occurred in late leptotene and
non-destructive approach to induce unreduced female
pachytene stages of MMC, as evidenced by a relatively
gametes in P. alba x P. glandulosa. We will attempt to
higher percentage of triploids in the progeny, when com-
correlate meiosis in pollen mother cells (PMCs) with
pared to progeny from colchincine application at other
male bud phenological development and in turn corre-
meiotic stages. The present results demonstrated that
late male bud phenology with MMCs meiosis. We will
male bud phenology can be used to guide colchicine
then study the effects of colchicine treatments at differ-
applications to female buds and cause a more efficient
ent MMCs meiotic stages on ploidy, as indicated per-
Key words: chromosome doubling, effective treatment stage,female gamete, Populus alba x P. glandulosa, triploid. Material and Methods Introduction
In the present study, cuttings from sexually mature
Triploid plants are often characterized by properties of
trees of P. alba x P. glandulosa, which is native to Korea,
fast growth, large leaves and vigor in comparison with
and P. tomentosa, a timber species native to China,
their diploid counterparts. Natural or induced triploid
were. In early January 2004, floral branches were cut
aspen and poplars (Populus L.) can have certain growth
and hydroponically cultured in a greenhouse with day
and fiber characteristics that are superior to diploid
and night temperatures around 20 °C and 10 °C, respec-
forms (cf. SCHLARBAUM, 1991). The heterosis of triploids
in forest trees is usually the result of an increase inploidy level from fertilization between n gametes with
unreduced (2n) gametes (ZHU et al., 1995). In tree breed-
The morphological stage of male and female catkins
ing, triploids have been induced by chromosome dou-
were observed after floral branches were cultured in the
bling of pollen in Populus tremula L., P. tremuloides
greenhouse. The ratio of unfolded catkin/bud were mea-
Michx., P. deltoides Bartr. (JOHNSSON and EKLUNDH,
sured every 12 h, from 5 to 8 days in greenhouse condi-
1940; WINTON, 1968), P. alba L. (MASHKINA et al., 1989),
Hevea brasiliensis Mull. (ZHENG et al., 1983), P. tomen-tosa Carr. x P. bolleana Lauche P. alba x P. glandulosa
(ZHANG and LI, 1992; KANG et al., 1999).
Observations of MMCs and PMCs meiosis were peri-
KANG and ZHU (1997) found that 2n pollen germinated
odically conducted after the female and male floral
later on stigma and pollen tube grew slower than that of
branches of P. alba x P. glandulosa were cut and cul-
n pollen in P. tomentosa Carr. and P. tomentosa x P. bol-
tured in greenhouse. After 14 h in culture, 3–4 floral
leana, which suggested that 2n pollen does not have
buds were collected from the female and male floral
competitive advantage over n pollen and therefore, has
branches every 3 h until the ninth day. The number of
less chance of participating in fertilization. In contrast,
floral buds collected was about 702. The collected female
fertilization of 2n female gametes by n pollen would
and male floral buds were fixed in FAA (18:1:1, ethy1
have the potential to create greater numbers of
alcohol: formaldehyde: acetic acid) and Carnoy (3:1ethy1 alcohol: acetic acid) respectively, then stored at
1) Key Laboratory for Genetics and Breeding of Forest Trees and
4 °C. Meiosis was observed in PMCs excised from
Ornamental Plants, Ministry of Education, Beijing Forestry
anthers of the fixed floral buds. The PMCs were
University, P. O. Box 118, 100083, Beijing, China.
squeezed out of the anthers into a drop of aceto-carmine
) Corresponding author: XIANGYANG KANG. Telephone: 86-10-
62336104, E-mail: [email protected].
Pistillate flowers at various developmental stages
were removed from catkins, dehydrated by consecutive
The ratio of triploids/seedlings was calculated in each
immersion in 70 %, 85 %, 95 % and 100 % ethanol (2 h for
colchicine treatment. The meiotic stage of MMCs was
each immersion), embedded in paraffin (MA et al, 2002)
determined by the percentage of MMCs meiotic stage in
sectioned between 8–10 µm, and dyed with 0.5 % hema-
each floral development. Pearson’s correlation coefficient
toxylin. The preparations were observed under an Olym-
was used to evaluate correlation between the ratio of
pus BX51 microscope and photographed. At least 3–4
triploids/seedlings and the percentage of MMCs meiotic
buds were sampled and 50–60 ovules were checked for
stage. Statistical analysis were performed by one-way
analysis of correlation coefficient using Data processingsystem 7.55 software(DPS7.55). Statiscial significant
Chromosome doubling of female gametes
difference was assumed when P value was < 0.05.
After associating MMCs and PMCs meiotic stages
with male bud development, eight female floral buds of
P. alba x P. glandulosa were immersed in a 0.5 % solu-
Association of MMCs and PMCs of P. alba x P. glandu-
tion of colchicine (LI et al., 2001) for 24 h with the excep-
tion of the controls (four female floral buds). Colchicinetreatments were initiated at 140 h, 152 h, 164 h, 176 h
Cytological observations of MMCs and PMCs meiosis
and 188 h respectively after the floral branches were
revealed that there was a close association betweenMMCs meiotic stage and male bud phenology during
cultured in greenhouse ranging from tetrad to two celled
development (Table 1). After 140 h in culture, when the
pollen stage of PMCs during each meiotic stage of
meiosis stage of PMCs developed into tetrads, the meio-
MMCs (leptotene, late leptotene, pachytene, diplotene,
sis stage of MMCs of P. alba x P. glandulosa was just
diakinesis and metaphase I). Pollination was conducted
beginning leptotene, and 1/4 pistillate catkin and 1/2
with P. tomentosa when the stigma was at the optimal
staminate catkin were protruded from squama respec-
receptive stage, as evidenced by a large amount of secre-
tively. After 164 h in culture, the meiosis stage of MMCs
tion After pollination, the female floral branches were
was in pachytene stage, and 3/4 staminate catkin and
further cultured in water until seeds matured and were
1/2 pistillate catkin were protruded from squama
harvested, approximately 2–3 weeks. Subsequently the
respectively. Within the female buds in a pistillate
seeds were sown in soil for each treatment and control.
catkin, most MMCs were synchronized meiotically.
After 2 months the seedlings were transplanted to field.
Table1 also showed the association between the morpho-logical characteristics of catkins and the stage of
Chromosome counts of two-year-old hybrid progenies
were determined using shoot tips prepared according to
the protocol developed by KANG (1996) and stained with
Table 2 presents the results obtained in terms of num-
ber of seedlings and percentage of triploid poplars
Table 1. – Association of MMCs and PMCs of P. alba x P. glandulosa.
a Hours after culture indicates the collection time of female buds. b unfolded catkin/bud indicates the ratio of unfolded catkin/bud. Figure 1. – Chromosomes of triploid poplars. (2n = 3x = 57). Figure 2. – Chromosomes of diploid hybrid progeny. (2n = 2x = 38).
among the hybrid progenies. A total of 12 seedlings were
percentage of triploids were obtained when the meiosis
identified as triploid poplars among 354 colchicine treat-
stage of most MMCs developed into late leptotene and
ment hybrid progenies (Figure 1), while the remaining
seedlings were diploid (Figure 2). Triploids were
Statistical analysis indicated that the rate of triploids
obtained in every colchicine treatment. There were no
was positive correlated with the late leptotene and
triploids obtained in control treatment. Guided by the
pachytene stage of MMCs meiotic stage, especially in
association between MMCs meiotic stage and male bud
diplotene stage, while the rate of triploids had negative
phenology during development, the female buds were
correlation coefficient with the leptotene, diakinesis and
treated with colchicine solution at the meiotic stage of
metaphase I stage of MMCs meiotic stage. There was
late leptotene (100 % MMCs). Two triploids were identi-
not statistically significant correlation between the rate
fied, accounting for 16.7% of the 12 survived two-year-
of triploids and percentage of MMCs meiotic stage
old seedlings. When treatment occurred at pachytene
(61.5 % MMCs) two triploid poplars were producedamong 87 seedlings total. Later pachytene treatments(75 % MMCs) produced three triploid poplars among 25
Discussion
seedlings total. Treatment at diakinesis (64.7 % MMCs)
The high rate of triploids was primarily obtained in
produced four triploids, which represented 5.0 % of 80
MMC late leptotene and pachytene stages and were
hybrid progenies. Overall, Table 2 shows that the high
associated with PMC and male bud phenology develop-
Table 2. – Triploid induction in P. alba x P. glandulosa by the chromosome doubling of female gamete.
c Hours after culture indicates the collection time of female buds and the treatment time of female buds. d Control: no colchicine treatment.
ment of P. alba x P. glandulosa. Although not statistical-
KANG, X. Y. and Z. T. ZHU (1997): A study on the 2n pollen
ly significant, the correlation trends suggests that a rel-
vitality and germinant characteristics of white poplars.
atively higher frequency of unreduced 2n female
Acta Botanica Yunnanica 19 (4): 402–406.
gametes were induced in late leptotene and pachytene.
KANG, X. Y. (1996): Chromosome count and shape of
It may be the induction of triploidy is so low that the
poplar. Journal of Gansu Agricultural University 31(1): 67–70.
statistical procedures used in this study would not show
KANG, X. Y., Z. T. ZHU and H. B. LIN (1999): Study on the
significance at the common error level of 0.05. Future
effective treating period for pollen chromosome dou-
studies that generate large number of seeds are needed
bling of Populus tomentosa x P. bolleana. Scientia Silvae
Sinicae 35(4): 21–24.
The high percentage of triploids (16.7 %) obtained in
KANG, X. Y., Z. T. ZHU and H. B. LIN (2000): Radiosensitiv-
the present study was higher than that of triploids
ity of different ploidy pollen in poplar and its applica-
(3.8 %) induced by the unreduced pollen in Populus
tion. Acta Genetica Sinica 27(1): 78–82.
ANG et al., 2000). Therefore, colchicine induction of 2n
I, Y., Z. T. ZHU, Y. T. TIAN, Z. Y. ZHANG and X. Y. KANG
female gametes using the association of MMCs with
(2001): Studies on obtaining triploids by colchicinetreating female flower buds of white poplars. Scientia
male bud phenology as a guide appears to be an efficient
Silvae Sinicae 37(5): 68–74.
approach for creation of triploids in P. alba x P. glandu-
MASHKINA, O. S., L. M. BURDAEVA, M. M. BELOZEROVA and
losa. Moreover, this approach avoids subjection of
L. N. V’YUNOVA (1989): A method of inducing diploid
female floral buds to high temperatures and daily appli-
pollen in woody species. Lesovedenie 1: 19–25 (in Russ-
cations of colchicine used in previous studies (LI et al,
MA, S.-M., X.-L. YE, N.-X. ZHAO and C.-Y. LIANG (2002):
Embryological studies on facultative apomixis in apluda mutica (Gramineae). Acta Botanica Sinica 44(3): Acknowledgements
SCHLARBAUM, S. E. (1991): Cytogenetics of forest tree
The authors thank Dr. G. HAZENBERG for critical read-
species. In: TSUCHIYA, T. and P. K. GUPTA, eds. Chromo-
ing of the manuscript. The research was financially sup-
some engineering in plant genetics and breeding. Vol.
ported by the State Forestry Administration of China
Elsevier Science Publ., Netherlands, p. 593–618.
(1999-12) and the National Natural Science Foundation of
WINTON, L. L. (1968): Fertilization in forced quaking
aspen and cottonwood. Silvae Genetica 17(1): 20–21.
ZHANG, Z. Y. and F. L. LI (1992): Studies on chromosome
doubling and triploid breeding of white poplar (_)-The
References
techniques of the pollen chromosome doubling, Journal of Beijing Forestry University, vol. 14, Supp. 3: 52–5.
DWIVEDI, N. K., N. SURYANARAYANA, A. K. SIKDAR, B. N.
ZHENG, X. Q., X. S. ZENG, X. M. CHEN and G. L. YANG
SUSHEELAMMA and M. S. JOLLY (1989): Cytomorphologi-
(1983): A new method for inducing triploid of Hevea
cal studies in triploid mulberry evolved by diploidiza-
brasiliensis. Chinese Journal of Tropical Crops 4(1):
tion of female gamete cells. Cytologia 54: 13–19.
JOHNSSON, H. and C. EKLUNDH (1940): Colchicine treat-
ZHU, Z. T., H. B. LIN and X. Y. KANG (1995): Studies on all
ment as a method in breeding hardwood species.
triploid breeding of Populus tomentosa B301 clones.
Svensk Papp Tidn 43: 373–377.
Scientia Silvae Sinicae 31(6): 499–505.
2013-14 Influenza Vaccine Registration and Consent VACCINE ADMINISTRATION RECORD Information about person who will receive vaccine (please print) Last Name First Name Birthdate MMDDYYYY Address: Street Phone Number Previous / Maiden name Clinic site: Heritage High School, Vancouver, WA I have been given a copy and have read or have had explained to me
Mind Cure, Robert A. Kloner, John Wiley & Sons, Incorporated, 1998, 0962602051, 9780962602054, . Dr. David Myers, world famous heart specialist, loved by his patients and peers, is losing his faculties and his job. Only a medical breakthrough can cure him and restore him to his former mental brilliance. But will a crazedcriminal seeking revenge interfere with the Mind Cure?. The Waiting Roo
 Pistillate flowers at various developmental stages
were removed from catkins, dehydrated by consecutive
The ratio of triploids/seedlings was calculated in each
immersion in 70 %, 85 %, 95 % and 100 % ethanol (2 h for
colchicine treatment. The meiotic stage of MMCs was
each immersion), embedded in paraffin (MA et al, 2002)
determined by the percentage of MMCs meiotic stage in
sectioned between 8–10 µm, and dyed with 0.5 % hema-
each floral development. Pearson’s correlation coefficient
toxylin. The preparations were observed under an Olym-
was used to evaluate correlation between the ratio of
pus BX51 microscope and photographed. At least 3–4
triploids/seedlings and the percentage of MMCs meiotic
buds were sampled and 50–60 ovules were checked for
stage. Statistical analysis were performed by one-way
analysis of correlation coefficient using Data processingsystem 7.55 software(DPS7.55). Statiscial significant
Chromosome doubling of female gametes
difference was assumed when P value was < 0.05.
After associating MMCs and PMCs meiotic stages
with male bud development, eight female floral buds of
P. alba x P. glandulosa were immersed in a 0.5 % solu-
Association of MMCs and PMCs of P. alba x P. glandu-
tion of colchicine (LI et al., 2001) for 24 h with the excep-
tion of the controls (four female floral buds). Colchicinetreatments were initiated at 140 h, 152 h, 164 h, 176 h
Cytological observations of MMCs and PMCs meiosis
and 188 h respectively after the floral branches were
revealed that there was a close association betweenMMCs meiotic stage and male bud phenology during
cultured in greenhouse ranging from tetrad to two celled
development (Table 1). After 140 h in culture, when the
pollen stage of PMCs during each meiotic stage of
meiosis stage of PMCs developed into tetrads, the meio-
MMCs (leptotene, late leptotene, pachytene, diplotene,
sis stage of MMCs of P. alba x P. glandulosa was just
diakinesis and metaphase I). Pollination was conducted
beginning leptotene, and 1/4 pistillate catkin and 1/2
with P. tomentosa when the stigma was at the optimal
staminate catkin were protruded from squama respec-
receptive stage, as evidenced by a large amount of secre-
tively. After 164 h in culture, the meiosis stage of MMCs
tion After pollination, the female floral branches were
was in pachytene stage, and 3/4 staminate catkin and
further cultured in water until seeds matured and were
1/2 pistillate catkin were protruded from squama
harvested, approximately 2–3 weeks. Subsequently the
respectively. Within the female buds in a pistillate
seeds were sown in soil for each treatment and control.
Pistillate flowers at various developmental stages
were removed from catkins, dehydrated by consecutive
The ratio of triploids/seedlings was calculated in each
immersion in 70 %, 85 %, 95 % and 100 % ethanol (2 h for
colchicine treatment. The meiotic stage of MMCs was
each immersion), embedded in paraffin (MA et al, 2002)
determined by the percentage of MMCs meiotic stage in
sectioned between 8–10 µm, and dyed with 0.5 % hema-
each floral development. Pearson’s correlation coefficient
toxylin. The preparations were observed under an Olym-
was used to evaluate correlation between the ratio of
pus BX51 microscope and photographed. At least 3–4
triploids/seedlings and the percentage of MMCs meiotic
buds were sampled and 50–60 ovules were checked for
stage. Statistical analysis were performed by one-way
analysis of correlation coefficient using Data processingsystem 7.55 software(DPS7.55). Statiscial significant
Chromosome doubling of female gametes
difference was assumed when P value was < 0.05.
After associating MMCs and PMCs meiotic stages
with male bud development, eight female floral buds of
P. alba x P. glandulosa were immersed in a 0.5 % solu-
Association of MMCs and PMCs of P. alba x P. glandu-
tion of colchicine (LI et al., 2001) for 24 h with the excep-
tion of the controls (four female floral buds). Colchicinetreatments were initiated at 140 h, 152 h, 164 h, 176 h
Cytological observations of MMCs and PMCs meiosis
and 188 h respectively after the floral branches were
revealed that there was a close association betweenMMCs meiotic stage and male bud phenology during
cultured in greenhouse ranging from tetrad to two celled
development (Table 1). After 140 h in culture, when the
pollen stage of PMCs during each meiotic stage of
meiosis stage of PMCs developed into tetrads, the meio-
MMCs (leptotene, late leptotene, pachytene, diplotene,
sis stage of MMCs of P. alba x P. glandulosa was just
diakinesis and metaphase I). Pollination was conducted
beginning leptotene, and 1/4 pistillate catkin and 1/2
with P. tomentosa when the stigma was at the optimal
staminate catkin were protruded from squama respec-
receptive stage, as evidenced by a large amount of secre-
tively. After 164 h in culture, the meiosis stage of MMCs
tion After pollination, the female floral branches were
was in pachytene stage, and 3/4 staminate catkin and
further cultured in water until seeds matured and were
1/2 pistillate catkin were protruded from squama
harvested, approximately 2–3 weeks. Subsequently the
respectively. Within the female buds in a pistillate
seeds were sown in soil for each treatment and control.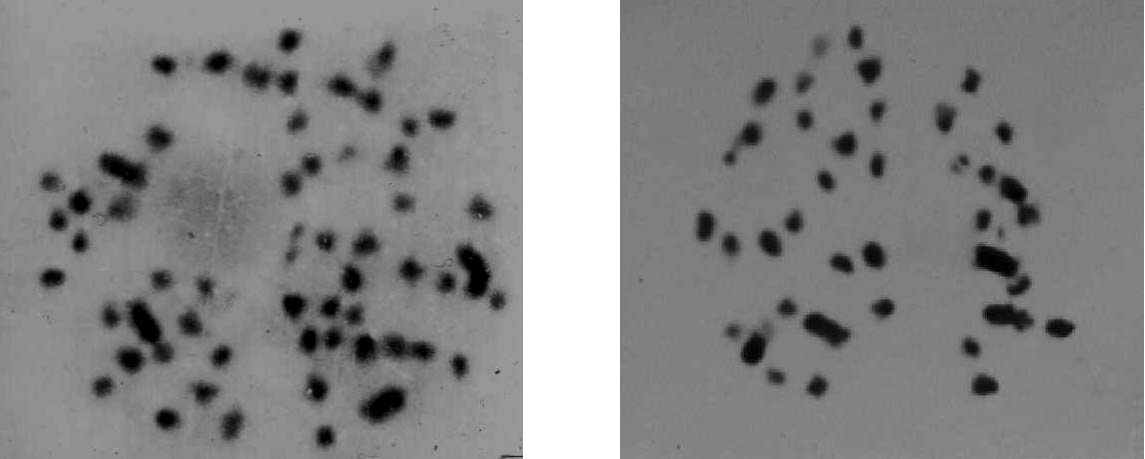
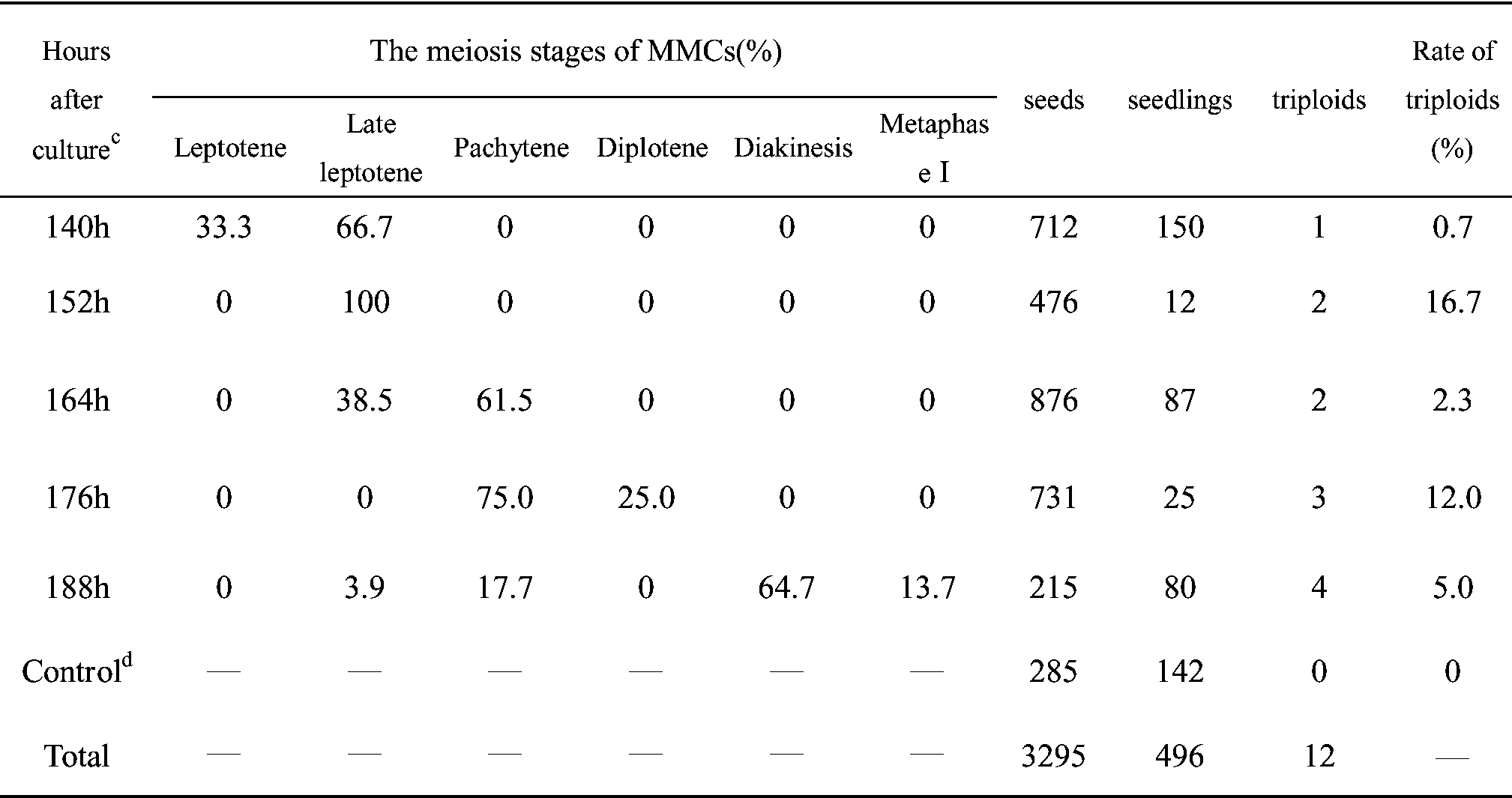 Figure 1. – Chromosomes of triploid poplars. (2n = 3x = 57).
Figure 1. – Chromosomes of triploid poplars. (2n = 3x = 57).