Age-specific mortality analysis of the dry forest kissingbug, Rhodnius neglectus
Jorge E. Rabinovich1*, Eliana L. Nieves1 & Luis F. Chaves21Centro de Estudios Parasitolo´gicos y de Vectores, Universidad Nacional de La Plata, La Plata, Prov. de Buenos Aires, Argen-tina, and 2Department of Environmental Studies, Emory University, Atlanta, GA, USA
Key words: senescence, cohort studies, Chagas disease, mortality rate, Weibull model, Gompertzmodel, exponential model, logistic model, DeMoivre model, Heteroptera, Reduviidae
Age-specific mortality patterns can be very different across insects with different life histories. Someholometabolous insects (like mosquitoes, fruit flies) show a pattern where mortality rate deceleratesat older ages, whereas other holometabolous insects (bruchid beetles) and hemimetabolous insects(cotton stainers, milkweed bugs, and kissing bugs) show an age-specific mortality pattern thatincreases through all ages. Kissing bugs are strictly hematophagous and are vectors of Trypanosomacruzi Chagas, the etiologic agent of Chagas disease. Here, we tested whether cohort data from the dryforest kissing bug, Rhodnius neglectus Lent (Hemiptera: Reduviidae), supports an increase of mortal-ity rate that decelerates with age. We analyzed the age-specific mortality pattern of a cohort of 250individuals of R. neglectus. We used a suite of seven models with different degrees of complexity, tomodel age-dependent forms of change in mortality rate increase in R. neglectus in the laboratory. Weused the Akaike model selection criterion to choose between models that consider absence or pres-ence of mortality deceleration. Five of the seven models (logistic, Gavrilovs, Gompertz, DeMoivre,and exponential) showed a statistically significant fit to the mortality rate. Weak late-age mortalitydeceleration in R. neglectus was supported by the best fit (logistic model), and this result is consistentwith predictions of the disposable soma theory of senescence.
with very different life histories. Holometabolous insects,
such as mosquitoes (Styer et al., 2007), fruit flies (Curt-
The understanding of senescence, that is, the increase in
singer et al., 1992; Carey et al., 1992, 2005; Fukui et al.,
mortality rate with age because a decline in physiological
1993), but not a bruchid beetle (Tatar et al., 1993), have
functioning, is a major goal of the research agenda in
shown a pattern where mortality decelerates at older ages,
evolutionary ecology (Williams, 1957; Abrams, 1993;
eventually reaching a plateau. Studies in hemimetabolous
Gavrilov & Gavrilova, 2001; Carey, 2003). The need to
insects, including the cotton stainer (Dingle, 1966), the
fully understand its underpinnings requires the use of the
milkweed bug (Dingle, 1966), and two species of kissing
comparative approach (Carey, 2001) which builds upon
bugs (Chaves et al., 2004a,b), have shown a pattern where
observations of a wide variety of organisms. Most of the
studies have been focused on a small subset of species, with
Kissing bugs have strict hematophagy across all of their
the most comprehensive studies coming from insects,
foraging ontogenetic stages, which is associated with a
especially fruit flies (e.g., Carey, 2003). It has been observed
mechanism that avoids the accumulation of oxidative sub-
that mortality patterns can be very different across insects
stances associated with aging (Grac¸a-Souza et al., 2006). They are also vectors of Trypanosoma cruzi Chagas, theetiologic agent of Chagas disease (WHO, 1991). Thus, the
*Correspondence: Jorge E. Rabinovich, Centro de Estudios Parasit-
study of senescence in this group may contribute to the
olo´gicos y de Vectores, Universidad Nacional de La Plata, Calle 2 No.
study of senescence as an evolutionary process and prove
584, 1900 La Plata, Prov. de Buenos Aires, Argentina. E-mail: jorge.ra-
useful for pest population management. Here, we present
a cohort study of the dry forest kissing bug, Rhodnius
Ó 2010 The Authors Entomologia Experimentalis et Applicata 1–11, 2010
Journal compilation Ó 2010 The Netherlands Entomological Society
neglectus Lent (Hemiptera: Reduviidae) focusing on the
serving both as resting place and for climbing to the top at
analysis of age-specific mortality and survivorship. As pre-
vious studies on kissing bugs (using Rhodnius prolixus Sta˚l
Each cohort was fed weekly using hens placed on a woo-
and Rhodnius robustus Larrousse) were focused on the
den box with holes at the bottom, through which the tops
Gompertz and the positive-shaped Weibull models
of the cohort jars could be tightly inserted. The insects
(Chaves et al., 2004a,b) we compare these two models,
climbed to the top and fed through the nylon mesh. Food
which cannot account for mortality rate deceleration, to
was offered for 1 h, and during the following hour the jars
an additional suite of five models with various degrees of
were horizontally exposed to a fan, to avoid the accumula-
complexity, that can model mortality rate deceleration and
tion of excessive moisture in the glass containers, which is
different forms of mortality rate increase with age (Appen-
common after the insects become engorged. The hens were
dix A). Our aim is to test whether the instantaneous mor-
sedated using an intramuscular dose of midazolam
tality rate in R. neglectus decelerates with age. We compare
(0.25 mg kg)1), a sedative commonly used to relax small
model fitting using Akaike Information Criterion model
animals before surgery. We allowed the hens to rest for
selection to test the absence or presence of instantaneous
5–10 min after the sedative application, and then carefully
rate of mortality deceleration in this species.
plucked them on one side on a surface not larger than adiameter of 7 cm (the jars’ mouth size). A towel was usedto restrain the hens to minimize stress from being
restrained. This procedure was carried out by two people
who would gently wrap the towel around the hen, letting
Rhodnius neglectus is a species that has been observed in
the head and legs stick out for better freedom of move-
Northeast Brazil (Lent & Wygodzinsky, 1979; Carcavallo
ments. This procedure was approved by the Animal Use
et al., 1999; Galvao et al., 2003) and in Venezuela in the
Institutional Review Board of IVIC (Venezuelan Institute
State of Amazonas, until at least 1965 (Gamboa Cuadrado,
for Scientific Research) and is in conformity with Venezu-
1973). This species is found in a region characterized by
mean annual temperatures ranging between 19 and 30 °C
After being exposed to the fan, each jar was opened to
(average 23.3 °C), and a mean (±SD) precipitation of
check the number of dead individuals (identified by
1 246 ± 429 mm year)1. Rhodnius neglectus has been
instar); identification by sex was performed only for
identified by Curto de Casas et al. (1999) to occupy the
adults, and only the female population was recorded.
Holdridge Dry Forest and Very Dry Forest or Savannah
Thus, data on mortality during the nymphal stages reflect
a mix of the two sexes, whereas the adult stage reflects onlyfemales. In two instances, the strict weekly feeding and
counting schedule could not be maintained (although
The population of R. neglectus used was made available by
they were never out of phase for more than 3 days), and
Dr Rodolfo Carcavallo and came from the insectary of the
as the biodemographic methods require a constant time
Instituto Oswaldo Cruz (Rio de Janeiro, Brazil), but its
unit for analysis, the recorded information was subjected
exact geographical origin is not known. The study was car-
to linear interpolation to keep the week as the time unit
ried out in Caracas, Venezuela, in a climate room with
for calculation of the age-specific mortality rates. Addi-
constant conditions of temperature (26 ± 1 °C) and
tionally, this procedure had to be applied at the beginning
humidity (60 ± 10% r.h.). Regulation of temperature is
of the cohort study (when the number of individuals was
essential because of the strong effect it has on the develop-
still high). This interpolation is a sound procedure,
mental cycle and survival (Garcia da Silva & da Silva,
because (1) when the cohorts are still in their first ages, all
1988). The ambient photoperiod was used: this did not
models predict an exponential mortality, which for small
need to be controlled, because at the latitude of Caracas its
x values can be considered linear (due to the Taylor
seasonal variation is very small (day length in December is
expansion; Demidovich, 1973), and (2) the various mod-
11:29 hours, and in June 12:42 hours).
els do not discriminate mortality patterns at early ages of
The design involved the follow up of five independent
the cohort, that is, most of them coincide in their predic-
cohorts initiated simultaneously. Each cohort was started
tions for early ages, yet they can predict very different pat-
with 100 recently laid (0- to 48-h old) eggs, kept in 150-ml
glass containers until all viable eggs hatched. The firstinstars were transferred to 3.8-l jars, covered with nylon
mesh, and with vertically placed strips of paper inside,
The weekly recording of the numbers of bugs alive anddead was used to calculate, following Carey (2001), the
basic parameters of a decrement life table: (1) survival as a
well as a description of the models and parameters used,
function of age (lx = Nx ⁄ N0, or fraction alive at age x), (2)
age-specific (period) survival (px = lx+1 ⁄ lx, or fractionalive at age x surviving to x + 1), (3) age-specific (period)
mortality (qx = 1 – px, or fraction alive at x dying prior to
Carey et al. (1992) claim that survival curves are poorly
suited for summarizing age-specific mortality patterns as
The age-specific mortality, qx, is a discrete quantity, rep-
compared with mortality rate curves, so we fitted models
resenting mortality as the probability of dying over the
to data using lx. For that purpose, the data used to fit the
1-week interval used. The continuous analog of this mea-
models were converted from their initially lx format, into
sure is referred to as the force of mortality or instanta-
px = lx+1 ⁄ lx, and then to mortality rates by lx = -ln(px).
neous mortality rate and denoted as l(x); it is defined as
All models were fitted using various procedures in the R
the mortality rate representing the limiting value of the
language (R Development Core Team, 2007). Because of
age-specific mortality rate when the age interval to which
the small number of mortality rate observations between
the rate refers becomes infinitesimally short (Carey, 2001).
weeks 83 and 89, when the last bugs died, we only analyzed
The age-specific mortality rate is preferred over age-spe-
the first 83 weeks, because estimates of the mortality rate
cific mortality, qx, because it is not bounded by unity, it is
(lx) become unreliable given the small number of individ-
independent of the size of the age intervals, and it is used
uals that can face death (Carey et al., 1992). Despite the
in numerous mortality models (Carey, 2001). The analyti-
number of individuals decreasing with age, we did not
cal relationship between the discrete form of mortality, qx,
weight the data because the models used are non-linear
and its continuous expression is l(x) = )ln px and
regression models that constitute an alternative to a weigh-
ing function in linear regression analyses. The resulting
We first analyzed the five cohorts independently, and
time series was analyzed both raw and smoothed; the rea-
carried out a Friedman ANOVA and a Kendall coefficient
son for smoothing before model fitting is that the data
of concordance test to verify if the differences in mortality
become easier to handle by the models by removing irreg-
rates l(x) among cohorts were statistically significant.
ularities and inconsistencies, thus capturing better the
Although mortality rates in older ages were not synchro-
behavior of the mortality rate process being modeled
nized, results showed that l(x) differences among
(Carey, 2001). For smoothing we used the Kolmogorov–
the five cohorts were not statistically significant
Zurbenko adaptive filter (KZA) (Chaves et al., 2008), with
(ANOVA: v2 = 5.902, P = 0.21; Coefficient of concor-
parameter q = 2 (half the size of the smoothing window).
dance = 0.0168, Average rank r = 0.00547). The further
The exponential model was fitted using least squares (Far-
analysis of the age-specific mortality pattern was carried
away, 2005) and confidence intervals obtained from the
out after pooling the five cohorts, treating the data as a
standard errors. For this model no differences were
single cohort. The initial number of eggs in the pooled
observed when compared with a weighted regression, with
data was taken 250, because the average sex ratio was 0.44
weights proportional to the number of surviving individu-
[$ ⁄ ($+#)] (which is not significantly different from 0.5;
als. The Gavrilovs model was fitted using the Nelder–Mead
two-sided t-test: P = 0.18; StatSoft, 2009). Similarly,
optimization algorithm (Nelder & Mead, 1965) imple-
unbiased 0.5 sex ratios have been found in other triato-
mented with the R command ‘optim’, and 95% confidence
mine species (Ronderos, 1972; Paz Rodrı´guez, 1996).
intervals were found using a non-parametric bootstrap
Despite the five cohorts being pooled for their fitting to
(Faraway, 2005) on 1 000 replications. All other models
mortality rate models, the results of the individual
were fitted by non-linear least squares (Bates & Watts,
cohorts are also presented to provide visualization of the
1988), using the R command ‘nls’. For these models, SEs
dispersion of the data of the cohorts around their pooled
were computed with a non-parametric bootstrap similar
to the one described for the Gavrilovs model.
In the analysis of the age-specific mortality pattern of
the pool of the five cohorts, the following six mortality
models were used: the exponential, deMoivre, Gompertz,
To compare the performance of different models with var-
Gompertz-Makeham, Weibull, and logistic models, using
ious degrees of freedom (i.e., different number of fitted
the algebraic presentation by Carey (2001), and a seventh
parameters), we used the Akaike Criterion (AIC) (Akaike,
three-parameter model based on reliability theory as pro-
1974), a method preferred over classical goodness-of-fit
posed by Gavrilov & Gavrilova (2001); for simplicity the
tests (such as the chi-squared or the G-tests) because it
latter will be called hereafter ‘Gavrilovs’. The reasons for
‘penalizes’ models with a high number of parameters. The
using a mortality analysis and not a survival analysis, as
AIC evaluation of model performance (one of the most
used goodness-of-fit indicators) represents the ‘informa-tion content’ of a given set of parameter estimates (Burn-ham & Anderson, 2002), and it is independent of themagnitude of the data points and of the number of obser-vations. It is calculated as:
where Yobsi and Ycalci are the observed and predicted mor-tality rates for the ith observation, ‘p’ is the number ofparameters, and ‘n’ the number of observations. The smal-ler the value of the AIC, the more appropriate the model is.
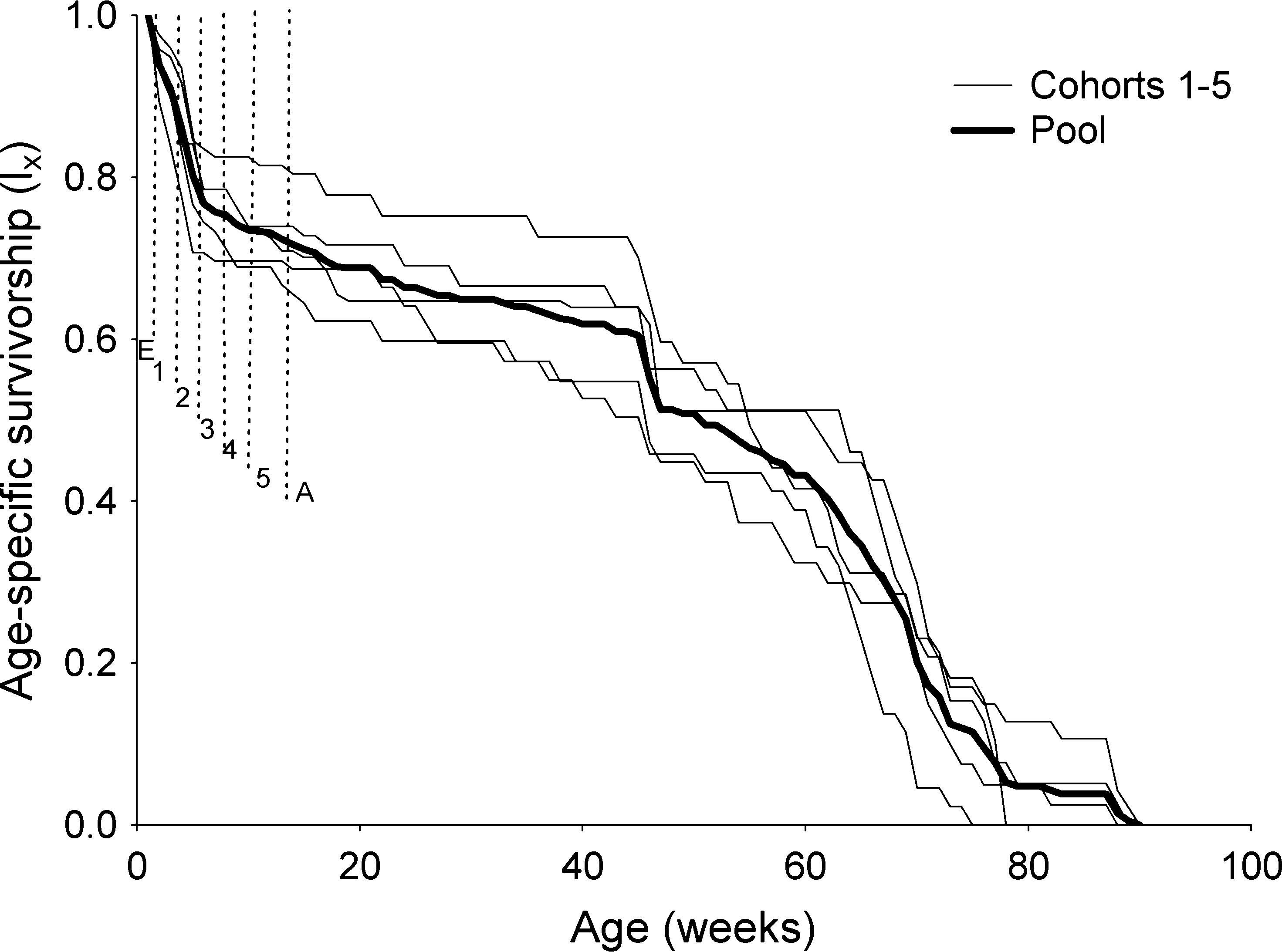
Figure 1 Age-specific survival (lx) for each cohort of Rhodnius
After converting the cohort age-specific survival (l
neglectus and the pool of the five cohorts. Vertical dashed lines
indicate the average periods of immature development. E, eggs;
x and l(x), and despite the peaks of mortality in later ages
1–5, instars 1–5; A, female adults.
not being synchronized, the differences between theage-specific mortality rate l(x) among the five cohorts ofR. neglectus are not statistically significant (Table 1),indicating that the five cohorts may be pooled.
The age-specific female survival (lx) values for each
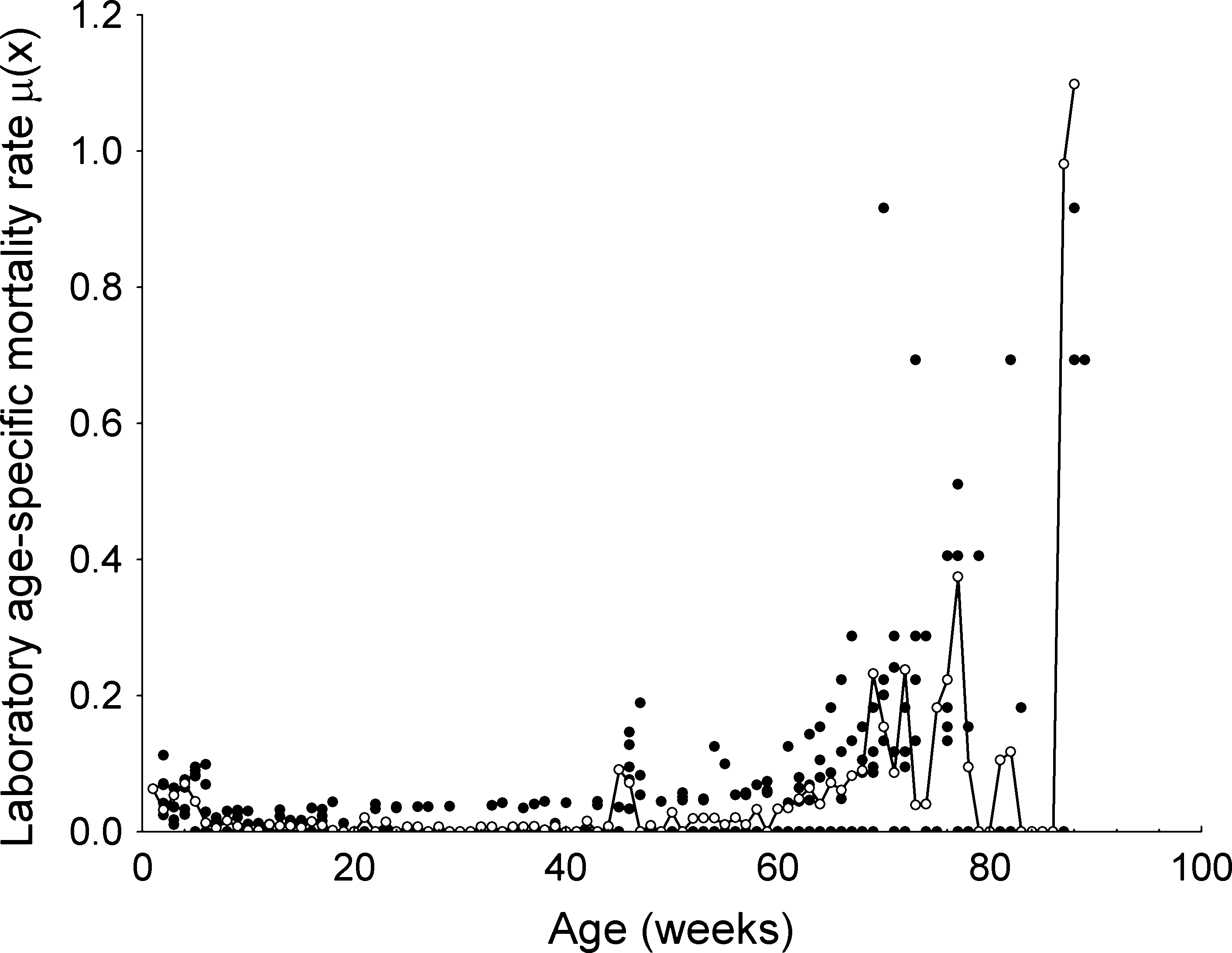
cohort as well as the pool of the five cohorts are presentedin Figure 1. There is a sharp decline in survival in the firstdevelopmental stages (egg, and instars 1 and 2), then thedecline stabilizes until approximately the age of 45 weeks(about 30 weeks of adult female’s age) when another sharpdecline is observed; after age 45 there is a progressivedecline in survival. Figure 2 shows the weekly evolution ofthe age-specific mortality rate l(x) for the individualcohorts and for the pool among cohorts.
Some of the results of the fitting of the smoothed and
non-smoothed data to the models are shown in Figure 3and Table 2. The Weibull model is not shown graphicallybecause it had a statistically non-significant fit to one of its
Figure 2 Age-specific mortality rate [l(x)] patterns of each
parameters. Figure 3 shows the predictions of the remain-
cohort of Rhodnius neglectus (black dots) and the pool of the fivecohorts (white dots and solid line). The individual cohorts were
ing five models (Gompertz, exponential, DeMoivre, Gavri-
not identified separately to facilitate visibility.
lovs, and logistic) with observed pooled laboratory data of
Table 1 Friedman ANOVA and Kendall coefficient of concor-
the age-specific mortality rate l(x) of R. neglectus. The
dance to test the differences in the weekly mortality rates [l(x)]
parameter values of those five age-specific mortality rate
sequence among the five cohorts of Rhodnius neglectus
models tested are shown in Table 2 for the raw data andthe KZA smoothing. Figure 4 shows the maximum effort
of egg laying. The quadratic fit for fecundity, given by the
equation y = -0.0085x2 + 0.7386x - 5.4479 (r = 0.908), is
shown only to ease visualization of the peak of the repro-
ductive effort of R. neglectus females. The method for
calculating fecundity (eggs ⁄ female ⁄ week) is given in Rabi-
novich (1972). It is striking how the peak of reproductive
ANOVA v2 = 5.902, d.f. = 4, P = 0.21; n = 88.
effort of R. neglectus coincides with the second early peak
Coefficient of concordance = 0.0168, average rank r = 0.00547.
In recent years, some striking lack of fit to the Gompertzmodel has been found in large cohorts of experimentalanimals, primarily insects (Partridge & Mangel, 1999). Theresults of our study show weak evidence for late-age mor-tality deceleration in R. neglectus, in contrast to patternsreported for several Diptera, including the fruitfly Dro-sophila melanogaster Meigen (Curtsinger et al., 1992;Pletcher & Curtsinger, 1998), the medfly Ceratitis capitata(Wiedemann) (Carey et al., 1992, 1998), the mexfly Ana-strepha ludens (Lo¨w) (Carey et al., 2005), and the mosquitoAedes aegypti (L.) (Styer et al., 2007). Rhodnius neglectusage-specific mortality pattern seems to be more similar tothat reported for other kissing bugs such as: R. prolixus
Figure 3 Age-specific mortality rate l(x) of observed laboratorydata (as pool of five cohorts) of Rhodnius neglectus and the pre-
(Chaves et al., 2004a), R. robustus (Chaves et al., 2004b),
dictions of the DeMoivre, exponential, Gompertz, logistic, and
the milkweed bug, Oncopeltus fasciatus (Dallas) (Dingle,
Gavrilovs models, for the smoothed data applied to l(x).
1966), the cotton stainer, Dysdercus fasciatus Signoret(Dingle, 1966), and the bruchid beetle Callosobruchusmaculatus (Fabricius) (Tatar et al., 1993). This result is
To determine the model that represents best the age-
consistent with the equilibrium predictions of the antago-
specific mortality rate data of R. neglectus, the results of the
nistic pleiotropy and mutation accumulation models of
application of the Akaike model selection criterion to the
senescence (decline in physiological functioning with age;
seven mortality rate models are shown in Table 3 for the
Abrams & Ludwig, 1995). These models are known as the
KZA smoothed l(x) data. From the models that con-
‘disposable soma’ theory for the evolution of senescence
formed significantly to the data (the Weibull and GM
(Kirkwood & Rose, 1991). This theory states that senes-
models did not have a significant fit), the logistic and the
cence arises from an optimal balancing of resources
Gompertz conformed best, and DeMoivre and exponential
between reproduction and somatic repair. For instance,
the worst. Similar results (not shown) were obtained with
model simulation showed that only when reproductive
output gradually approaches, but never reaches, a maxi-mum value as the amount of diverted resources increases,senescence is consistent with the Gompertz model(Abrams & Ludwig, 1995).
Table 2 Parameter values of the five age-specific mortality rate [l(x)] models (the Weibull and Gompertz-Makeham models were notincluded because their fit to the laboratory data was not statistically significant)
‘p’ is the number of parameters of each model. In brackets are the 95% confidence intervals; intervals that do not contain zero are statistic-ally significant with P<0.05, and have been presented graphically in Figure 3. KZA refers to the Kolmogorov–Zurbenko adaptive filtersmoothing method. The models are presented in increasing order of their number of parameters.
found in the beetle C. maculatus (Tatar et al., 1993) andthe kissing bug R. prolixus (Sulbaran & Chaves, 2006). Williams (1957) claimed that greater rates of ‘extrinsic’(age- and condition-independent) mortality favored morerapid senescence. However, Abrams (1991) showed thatthe effects of the ‘extrinsic’ mortality differentially affectthe rate of senescence according to density-dependence. Furthermore, Abrams (1991) showed that mortality pat-terns, contrary to Williams (1957) predictions, are possiblewhen density-dependence is present, and acts primarily onthe survival or fertility of later ages, or when most of thevariation in mortality rates is due to variation in non-extrinsic mortality. No experimental evaluation exists ondensity-dependent processes in R. neglectus. However,Rodrı´guez & Rabinovich (1980) showed in R. prolixus that
Figure 4 Age-specific survival of the pool of five cohorts of
density had a significant effect on the developmental rate
Rhodnius neglectus from the egg stage and the maternity function(eggs ⁄ female ⁄ week) as an indicator of egg laying effort. The
of second, third, and fourth instars, but not on first instars,
maternity function was fitted to a quadratic equation (y =
adult survivorship, instantaneous population parameters,
)0.0085x2 + 0.7386x - 5.4479, r = 0.908) only to ease visualiza-
or age-specific parameters. If similar patterns were present
tion of the peak of the reproductive effort of R. neglectus females.
in R. neglectus, then this would confirm that in this speciesWilliams’ (1957) rapid senescence may be a response to
Our results fit these predictions: we found a sharp
greater rates of ‘extrinsic’ mortality.
decline in survival in the early developmental stages (egg
In R. neglectus there is no apparent reduction of the
and instars 1 and 2), then the decline stabilizes at the age of
length of reproductive life as a consequence of high early
45 weeks (about 30 weeks of adult female’s age) when
fecundity (J.E. Rabinovich, unpubl.), unlike R. prolixus
another sharp decline is observed, possibly related to the
(Sulbaran & Chaves, 2006) and several strains of D. mela-
maximum effort of egg laying; after 45 weeks there is a
nogaster (Giesel & Zettler, 1980; Partridge, 2001). On the
progressive decline in survival. These two declines in l
contrary, R. neglectus shows a clear exponential increase in
correspond of course to the two strong earlier peaks of
death rate in older ages, suggesting a Gompertz behavior,
mortality rate l(x) as shown in Figure 3. Figure 4 further
and possibly reflecting a minor effect of the trade-off
supports the relationship between maximum effort of egg
between reproduction and somatic repair, confirming a
laying and the second early peak in the mortality rate. This
possible larger effect of ‘extrinsic’ mortality.
apparent relationship between egg-laying effort and effects
In contrast with the Gompertz model, in the Weibull
on the age-specific mortality rate is similar to the one
model the causes of death for young adults and old indi-viduals are different, independent, and additive, andinclude death causes due to catastrophic intrinsic sourceswith a probability that increases with age, and not as a
Table 3 Relative ranking (1 = best, 7 = worst) of the fit of seven
physiological function that declines with age as in the
mortality rate models to the KZA smoothed laboratory data of
Gompertz model (Gavrilov & Gavrilova, 2001; Ricklefs &
Rhodnius neglectus using Akaike model selection criterion (AIC)
Scheuerlein, 2002). Partly for these reasons, the Gompertzmodel is considered by most demographers as an empiri-
cal model rather than a ‘law’ (Carey, 2003). Despite the
Weibull function being usefully used in conjunction with
failure-time models (in which failure depends on the
occurrence of one or more rare events, such as genetic
mutations or cell deaths; Ricklefs & Scheuerlein, 2002), the
fit of this model to the R. neglectus mortality rate data was
The only other instance of application of mortality
Asterisks identify those models that had a statistically significant
models to kissing bug species are the studies of Chaves et
fit by the criterion of the 95% confidence intervals with the boot-
al. (2004a,b). Only the Gompertz model was tested on the
age-specific mortality rate of R. robustus, and the Gom-
pertz and Weibull models were tested on the age-specific
Triatoma infestans Klug, Triatoma brasiliensis Neiva, and
mortality rate of R. prolixus. The results from these two
Triatoma pseudomaculata Correia & Espinola, and Pan-
studies are different to those of R. neglectus in terms of the
strongylus megistus Burmeister was 37.5 and 45.9%, respec-
parameter values of both models. This may be partially
tively. The adaptation to a domestic environment causes a
due to demographic evolution of these taxa: R. prolixus has
similar effect on triatomines (Forattini, 1980) because
a population intrinsic rate of natural increase (rm) of about
conditions inside rural houses provide smaller predation
0.032 per week, almost double that of R. neglectus of 0.017
risks, smaller fluctuations of temperature and relative
per week (J.E. Rabinovich, unpubl.) reflecting their adap-
humidity that exclude deleterious extreme values, and bet-
tation as results of genetic selection to different environ-
ter chances of feeding. The values of the age-specific mor-
ments. This result also confirms the soundness of the
tality rate parameters estimated under domiciliary
assumptions of Charlesworth (2000), supporting the use
conditions will prove of importance in the epidemiology
of rm to explain the evolution of senescence and life-his-
of Chagas disease, and even decisive in the design of pest
The weak late-age mortality deceleration in R. neglectus
In a recent review of mammal and bird comparative
was observed only through the fit of the logistic model of
analyses of survival senescence by using life tables, Jones et
mortality, the model selected as ‘best’ by the Akaike model
al. (2008) have identified generalizations including the
selection criterion. However, this late-age mortality decel-
observation that mammals senesce faster than similar-
eration process is extremely sensitive to the sample size of
sized birds. Furthermore, McCoy & Gillooly (2008) devel-
the cohort study, and our initial cohort size of 250 individ-
oped a model of natural mortality (relating body size and
uals was probably too small to permit a strong assertion
temperature to biological rate processes) and tested it with
about late-age mortality deceleration in R. neglectus. As
extensive field data from plants, invertebrates, fish, birds,
claimed by Gaines & Denny (1993), when mortality rate is
and mammals; their results indicate that much of the het-
close to zero it behaves as a threshold character; and to esti-
erogeneity in natural mortality rates can be predicted,
mate the points ‘below threshold mortality’ becomes diffi-
explicitly and quantitatively, despite the many extrinsic
cult and their measures are usually relatively inaccurate
sources of mortality in natural systems, suggesting that
(Promislow et al., 1999). Carey (2003) considers that an
common rules govern mortality rates. We believe that
estimate of the threshold mortality can be taken as 1 ⁄ N,
comparative review is due for different orders of insects.
where N is the initial cohort size; in the case of R. neglectus
Such a review not only has academic value, but it would
the initial cohort size of 250 individuals would indicate a
also impinge on important pest population management
threshold mortality of 0.004, and mortality rate values
and epidemiological areas. New age-dependent models of
below this threshold exist in our laboratory data, both raw
pathogen transmission show the importance of age and
and smoothed. An increase in the initial cohort size is diffi-
population age structure to transmission dynamics (Styer
cult for kissing bugs, with a complex feeding behavior, the
et al., 2007). These authors claim that, in the case of mos-
need of large containers to avoid problems of overcrowd-
quitoes, a departure from the age-independent mortality
ing, and requirements of important numbers of adequate
paradigm is essential for an accurate understanding of
and live avian or mammal hosts for feeding.
mosquito biology and pathogen transmission.
However, we strongly suggest future studies with kissing
Environmental heterogeneity, and particularly high
bugs to consider this threshold mortality factor, which will
environmental uncertainty (stochastic effects) affects envi-
allow confirmation of whether hemimetabolous insects do
ronmental circumstances that are very important in
not decelerate their mortality at old ages, as our results
insects, such as encounter rates with suitable oviposition
suggest. As stated by Chaves et al. (2004a) in relation to
sites, food availability, physiological state (e.g., reserves for
R. prolixus, the mortality rate of this triatomine vector has
producing oocytes, egg maturation rate, and somatic
epidemiological importance through the demography of
maintenance costs), and expected reproductive success
its populations. In the case of R. neglectus, despite being
that can lead to different patterns of behavior and rates of
essentially sylvatic, it is considered to be in the process of
mortality and reproduction (Partridge & Mangel, 1999),
becoming a truly domestic species (Lent & Wygodzinsky
and they should be incorporated in future studies for
1979). It has been shown in triatomines that important
understanding senescence in insects.
changes in life-history traits occur when reared under lab-oratory conditions (Perlowagora-Szumlewicz, 1976); for
example, after only 4 years in the laboratory, the averagereduction in the number of eggs within the first month of
Data collection was carried out while the first author was a
oviposition and the average female life span among
researcher at the Instituto Venezolano de Investigaciones
Cientı´ficas, Caracas, Venezuela. We thank Diego Rodrı´-
Chaves LF, Zamora E & Aldana E (2004b) Mortality profile of
guez for helpful comments on an earlier draft of this man-
female Rhodnius robustus (Heteroptera: Reduviidae). Revista
uscript, and two anonymous reviewers for many useful
suggestions. L.F.C. was supported by a Post-Doctoral
Chaves LF, Kaneko A, Taleo G, Pascual M & Wilson ML (2008)
Fellowship from Emory University and a Gorgas Research
Malaria transmission pattern resilience to climatic variability ismediated by insecticide-treated nets. Malaria Journal 7: 100–
Award from the American Society of Tropical Medicine
and Hygiene. This manuscript benefited from discussions
Curto de Casas SI, Carcavallo RU, Galı´ndez Giro´n I & Burgos JJ
with several colleagues from a NIH-RAPIDD study group
(1999) Bioclimatic factors and zones of life. Atlas of Chagas’
Disease Vectors in the Americas, Vol. III. (ed. by RU Carcaval-lo, I Galı´ndez Giro´n, J Jurberg & H Lent), pp. 793–838. Edito-rial Fiocruz, Rio de Janeiro, Brazil.
Curtsinger JW, Fukui HH, Townsend DR & Vaupel JW (1992)
Abrams PA (1991) The fitness costs of senescence: the evolution-
Demography of genotypes: failure of the limited life-span para-
ary importance of events in early adult life. Evolutionary Ecol-
digm in Drosophila melanogaster. Science 258: 461–463.
Demidovich BP (1973) Problemas y Ejercicios de Ana´lisis
Abrams PA (1993) Does increased mortality favor the evolution
Matema´tico. Editorial Mir, Moscow, USSR.
of more rapid senescence? Evolution 47: 877–887.
Dingle H (1966) The effect of population density on mortality
Abrams PA & Ludwig D (1995) Optimality theory, Gompertz’
and sex ratio in the milkweed bug, Oncopeltus, and the cotton
law, and the disposable soma theory of senescence. Evolution
stainer, Dysdercus (Heteroptera). American Naturalist 100:
Akaike H (1974) A new look at statistical model identification.
Faraway JJ (2005) Linear Models with R. Chapman & Hall,
IEEE Transactions on Automatic Control 19: 716–723.
Bates DM & Watts DG (1988) Nonlinear Regression Analysis and
Forattini OP (1980) Biogeography, origin, and distribution of
its Applications. Wiley & Sons, New York, NY, USA.
triatominae domiciliarity in Brazil. Revista de Saude Pu´blica
Burnham KP & Anderson DR (2002) Model Selection and Multi-
model Inference. A Practical Information - Theoretical
Fukui HH, Xiu L & Curtsinger JW (1993) Slowing of age-specific
Approach. Springer, New York, NY, USA.
mortality rates in Drosophila melanogaster. Experimental Ger-
Carcavallo RU, Curto de Casas SI, Sherlock IA, Galı´ndez Giro´n I,
Jurberg J, et al. (1999) Geographical distribution and alti-lati-
Gaines SD & Denny MW (1993) The largest, smallest, highest,
tudinal dispersion of Triatominae. Atlas of Chagas’ Disease
lowest, longest, and shortest: extremes in ecology. Ecology 74:
Vectors in the Americas, Vol. III (ed. by RU Carcavallo, I Gal-
ı´ndez Giro´n, J Jurberg & H Lent), pp. 747–792. Editorial Fio-
Galvao C, Carcavallo RU, da Silva Rocha D & Jurberg J (2003) A
checklist of the current valid species of the subfamily Triatomi-
Carey JR (2001) Insect biodemography. Annual Review of Ento-
nae Jeannel, 1919 (Hemiptera, Reduviidae) and their geo-
graphical distribution, with nomenclatural and taxonomic
Carey JR (2003) Longevity. The Biology and Demography of Life
Span. Princeton University Press, Princeton, NJ, USA.
Gamboa Cuadrado J (1973) Distribucio´n geogra´fica y prevalencia
Carey JR, Liedo P, Orozco D & Vaupel JW (1992) Slowing of
de la poblacio´n de triatominos en Venezuela. World Health
mortality rates at older ages in large medfly cohorts. Science
Organization Planning Meeting for the Unit for the Study of
Vectors of Chagas Disease in Venezuela. Maracay 29–31
Carey JR, Liedo P, Mu¨ller H-G, Wang J-L & Vaupel JW (1998)
March, 1973. Direccio´n de Malariologı´a y Saneamiento Ambi-
Dual modes of aging in mediterranean fruit fly females. Science
Garcia da Silva I & da Silva HH (1988) Influencia da temperatura
Carey JR, Liedo P, Mu¨ller H-G, Wang J-L, Senturke D & Harsh-
na biologia de Triatomineos. II. Rhodnius neglectus Lent, 1954
manf L (2005) Biodemography of a long-lived tephritid:
(Hemiptera, Reduviidae). Revista Goiana de Medicina 34: 29–
Reproduction and longevity in a large cohort of female Mexi-
can fruit flies, Anastrepha ludens. Experimental Gerontology
Gavrilov LA & Gavrilova NS (2001) The reliability theory of aging
and longevity. Journal of Theoretical Biology 213: 527–545.
Charlesworth B (2000) Fisher, Medawar, Hamilton and the evo-
Giesel JT & Zettler EE (1980) Genetic correlations of life historical
lution of aging. Genetics 156: 927–931.
parameters and certain fitness indices in Drosophila melanogas-
Chaves LF, Herna´ndez MJ, Revilla TA, Rodrı´guez DJ & Rabino-
ter: rm, rs, diet breadth. Oecologia 47: 299–302.
vich JE (2004a) Mortality profiles of Rhodnius prolixus
Gompertz B (1825) On the nature of the function expressive of
(Heteroptera: Reduviidae), vector of Chagas disease. Acta Tro-
the law of human mortality and on a new mode of determining
life contingencies. Philosophical Transactions of the RoyalSociety, London 115: 513–585.
Grac¸a-Souza AV, Maya-Monteiro C, Paiva-Silva GO, Braz GR,
Rabinovich JE (1972) Vital statistics of Triatominae (Hemiptera:
Paes MS, et al. (2006) Adaptations against heme toxicity in
Reduviidae) under laboratory conditions. I. Triatoma infestans
blood-feeding arthropods. Insect Biochemistry and Molecular
Klug. Journal of Medical Entomology 9: 351–370.
Ricklefs RE & Scheuerlein A (2002) Biological implications of the
Jones OR, Gaillard J-M, Tuljapurkar S, Alho JS, Armitage KB,
Weibull and Gompertz models of aging. Journal of Gerontol-
et al. (2008) Senescence rates are determined by ranking on the
fast-slow life-history continuum. Ecology Letters 11: 664–673.
Rodrı´guez D & Rabinovich JE (1980) The effect of density on
Kirkwood TBL & Rose MR (1991) Evolution of senescence: late
some population parameters of Rhodnius prolixus (Hemiptera:
survival sacrificed for reproduction. Transactions of the Royal
Reduviidae) under laboratory conditions. Journal of Medical
Kohler H-P & Kohler I (2000) Frailty modelling for adult and old
Ronderos RA (1972) Biologı´a de Triatominos. Simposio Inter-
age mortality: the application of a modified Demoivre hazard
nacional sobre Enfermedad de Chagas, 26 de Noviembre al 2
function to sex differentials in mortality. Demographic
de Diciembre, pp. 321–326. Sociedad Argentina de Para-
sitologı´a, Buenos Aires, Argentina.
Lent H & Wygodzinsky P (1979) Revision of the Triatominae
StatSoft (2009) STATISTICA (Data Analysis Software System),
(Hemiptera, Reduviidae), and their significance as vectors of
version 9.0. Statsoft, Tulsa, OK, USA.
Chagas’ disease. Bulletin of the American Museum of Natural
Styer LM, Carey JR, Wang J-L & Scott TW (2007) Mosquitoes do
senesce: departure from the paradigm of constant mortality.
Makeham WM (1860) On the law of mortality and the construc-
American Journal of Tropical Medicine and Hygiene 76: 111–
tion of annuity tables. Journal of the Institute of Actuaries 6:
Sulbaran JE & Chaves LF (2006) Spatial complexity and the fit-
Makeham WM (1867) On the law of mortality. Journal of the
ness of the kissing bug, Rhodnius prolixus. Journal of Applied
Institute of Actuaries 13: 325–358.
McCoy MW & Gillooly JF (2008) Predicting natural mortality
Tatar M, Carey JR & Vaupel JW (1993) Long-term cost of repro-
rates of plants and animals. Ecology Letters 11: 710–716.
duction with and without accelerated senescence in Calloso-
Nelder JA & Mead R (1965) A simplex algorithm for function
minimization. The Computer Journal 7: 308–313.
Olshansky SJ & Carnes BA (1997) Ever since Gompertz. Demog-
Vaupel JW, Manton KG & Stallard E (1979) The impact of hetero-
geneity in individual frailty on the dynamics of mortality.
Partridge L (2001) Evolutionary theories of ageing applied to
long-lived organisms. Experimental Gerontology 36: 641–650.
WHO (1991) Control of Chagas disease. World Health Organiza-
Partridge L & Mangel M (1999) Messages from mortality: the
tion Technical Report Series 811. WHO, Geneva, Switzerland.
evolution of death rates in the old. Trends in Ecology & Evolu-
Williams GC (1957) Pleiotropy, natural selection and the evolu-
tion of senescence. Evolution 11: 398–411.
Paz Rodrı´guez R (1996) Ciclo de Vida y Datos Biome´tricos de
Triatoma longipennis (Usinger) (Hemiptera: Reduviidae,Triatominae). PhD Dissertation. Autonomous National
University of Mexico, Mexico City, Mexico.
Perlowagora-Szumlewicz A (1976) Laboratory colonies of
Triatominae, biology, and population dynamics. New
Approaches in American Trypanosomiasis Research. Pro-ceedings of an International Symposium, Belo Horizonte,
As stated in section ‘Statistical analysis’ under ‘Materials
Minas Gerais, Brazil, 18–21 March 1975, pp. 63–82. Pan
and methods’, two basic parameters of a decrement life
American Health Organization, Scientific Publications No.
table are directly related: survival as a function of age
(lx = Nx ⁄ N0), and age-specific mortality [qx = 1 –
Pletcher SD & Curtsinger JW (1998) Mortality plateaus and the
(lx+1 ⁄ lx)]. Thus we could have modeled survival (lx) or
evolution of senescence: why are old-age mortality rates so
mortality [qx; actually the instantaneous mortality rate,
denoted as l(x); that is, the rate representing the limiting
Promislow DEL, Tatar M, Pletcher S & Carey JR (1999) Below-
value of the age-specific mortality rate when the age inter-
threshold mortality: implications for studies in evolution,
val becomes infinitesimally short]. There are several rea-
ecology and demography. Journal of Evolutionary Biology 12:
sons why we preferred to use a mortality analysis and not
R Development Core Team (2007) R: A Language and Environ-
the method of survival analysis: (1) survival analysis is
ment for Statistical Computing. R Foundation for Statistical
based upon models like Kaplan–Meier or Cox, which esti-
Computing, Vienna, Austria. http: ⁄ ⁄ www.R-project.org ⁄
mate the death hazards but do not reflect any mechanismor process underlying the fitted curves as the models we
use do (survival models can be viewed as ordinary regres-
sion models in which the response variable is time), (2)
survival rates are mainly important for projections or
where lD(x) denotes the DeMoivre hazard function. The
prognosis, and to estimate the importance of cause-spe-
hazard rate increases towards infinity as x approaches the
cific survival (e.g., to rank the effects of different causes of
maximum attainable age x. This maximum age implies
mortality), which is not the goal of our analysis, (3) when
that the age-specific mortality rate increases faster than in
no truncation or censoring occurs, the Kaplan–Meier
the Gompertz model (see below), and in particular as x
curve is equivalent to the empirical distribution, and (4)
approaches x, when the mortality rate tends to approach
we were directly interested in the mortality and not the
In relation to the latter, despite the fact that mortality
and survival are intimately related, there is an important
Gompertz (1825), cited in Olshansky & Carnes, 1997) set
difference: death is an event whereas survival is a ‘non-
forth what is now recognized as the law of mortality. He
event’, that is, the absence of the mortality event (Carey,
noted that the differences between the common logarithm
2001). The advantage of the survival analysis method to
of the number of individuals living in successive equal age
deal with censored observations would not have improved
intervals were almost identical during a significant portion
our analysis, for in our study there were no censored
of their life span, in general at least after the age of sexual
observations (defining censored observations, as it is com-
maturity or another predetermined age, such as the maxi-
monly used in survival analysis, as those individuals who
mum reproductive effort (Carey, 2001, 2003). The mortal-
are lost to follow up being still alive or that remained alive
ity rate (lx) function in Gompertz’s model has the
when the study ended). Although we deleted observations
between weeks 83 and 89 because the mortality rate (lx)becomes unreliable given the small number of individuals
that can face death at those ages (see section ‘Fitting mod-
where ‘a’ is a parameter representing the initial mortality
els to data’ under ‘Materials and methods’), those observa-
rate, and ‘b’ is the Gompertz parameter that represents the
tions cannot be considered censored under our definition,
rate of increase of mortality with age (also called the Gom-
Makeham (1860) refined Gompertz’s law of mortality,
noting that the logarithms of the probabilities of living
This is the simplest mortality model that assumes that the
from Gompertz’s formula, instead of proceeding in uni-
mortality rate grows linearly with age x. The formula for
form geometrical progression, increased at a faster pace at
higher ages than at younger ages. He solved this problem
by adding a ‘constant’ term, and redefining the Gompertz
law as ‘the probabilities of living, increased or diminishedin a certain constant ratio, from a series whose logarithmsare in geometrical progression’. Later Makeham (1867)
modified Gompertz’s formula by developing ‘a theory of
The mortality rate in the DeMoivre model (1725, cited in
partial forces of mortality’. The Gompertz-Makeham
Kohler & Kohler, 2000) attempts to describe mathemati-
cally the mortality patterns assuming that ‘the number oflives existing at any age is proportional to the number of
years intercepted between the age given and the extremity
where ‘c’ is the additional parameter introduced by Make-
ham to represent the additional constant rate of mortality.
where x is the maximum attainable age in the population.
As with the Gompertz model, the Weibull model incorpo-
Then the hazard rate, or the age-specific mortality rate at
rates a minimum mortality rate suffered by young adults
prior to the onset of their physiological decline. This isusually referred to as the initial mortality rate (l0). After
this initial mortality the aging-related mortality rate
For this model mortality rates level off at older ages. This
increases exponentially as a multiple of the initial mortality
model is similar to a frailty model sensu Vaupel et al.
l0. In the Weibull model, the aging-related component of
mortality is a power function of age that is added to theinitial mortality rate:
Gavrilov & Gavrilova (2001) proposed a general model for
aging based on reliability principles. In this model it is
where ‘a’ and ‘n’ are the parameters of the Weibull model.
assumed that mortality rate [l(x)] comes from the deteri-
The value of ‘a’ determines the scale, and ‘n’ determines
oration of redundant subunits (n) organized in blocks (m)
the shape of the curve. The Weibull hazard function
that compose individuals, at a constant failure rate (k),
increases if n>0, decreases if n<0, and is constant if n = 0.
where the probability of an element being initially func-tional can vary from fully functional (q = 1) to highly
likely to be unfunctional (q $ 0). In this model the initial
The logistic model has a Gompertz-type mortality rate as a
number of functional elements has a binomial distribu-
baseline hazard, with hazard function (Carey, 2001):
Only the three-parameter version of this model was
tested (where m remains constant, i.e., m = 1).
EURAM-EDAMBA-EIASM DOCTORAL CONSORTIUM 2005 EURAM 2005 Pre-Conference Event Wednesday, May 4th 2005 (9:00 - 15:45 h) at the TUM Business School, City Campus, Munich, Germany, Room 1601 EURAM (European Academy of Management), EDAMBA (European Doctoral Programs Association in Management and Business Administration) and EIASM (European Institute for Advanced Studies in Management) invite
Brief History Mr J S was truly at his wits end. At 64, the ageing process had been far from beautiful and the years sat on him more like the proverbial Sword of Damoceles rather than an achievement of life. Eight years earlier, his coronary artery system had become disease afflicted. This resulted in a heart attack that severely curtailed his normal lifestyle. In order to remain physically
 used goodness-of-fit indicators) represents the ‘informa-tion content’ of a given set of parameter estimates (Burn-ham & Anderson, 2002), and it is independent of themagnitude of the data points and of the number of obser-vations. It is calculated as:
where Yobsi and Ycalci are the observed and predicted mor-tality rates for the ith observation, ‘p’ is the number ofparameters, and ‘n’ the number of observations. The smal-ler the value of the AIC, the more appropriate the model is.
used goodness-of-fit indicators) represents the ‘informa-tion content’ of a given set of parameter estimates (Burn-ham & Anderson, 2002), and it is independent of themagnitude of the data points and of the number of obser-vations. It is calculated as:
where Yobsi and Ycalci are the observed and predicted mor-tality rates for the ith observation, ‘p’ is the number ofparameters, and ‘n’ the number of observations. The smal-ler the value of the AIC, the more appropriate the model is.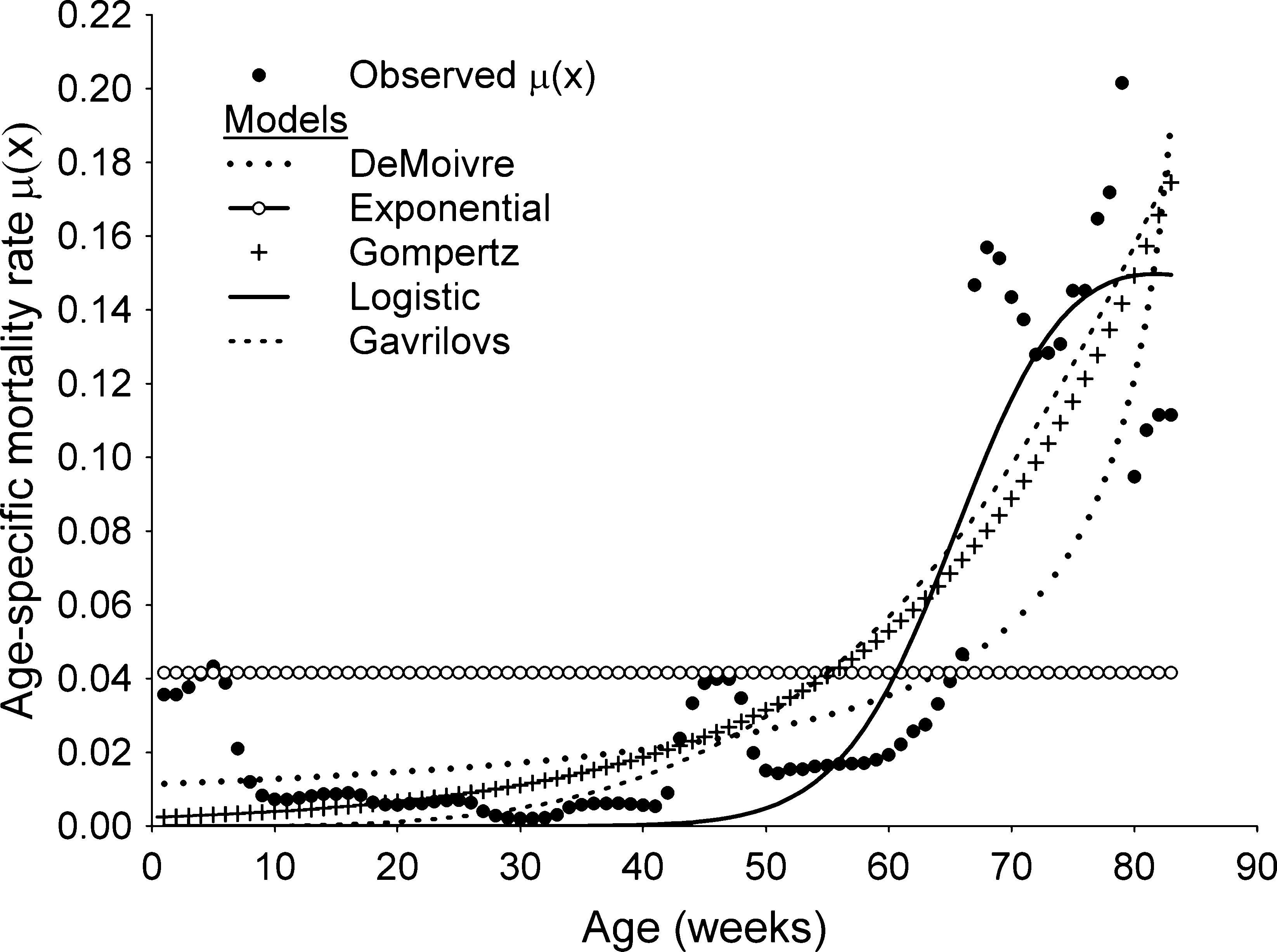 In recent years, some striking lack of fit to the Gompertzmodel has been found in large cohorts of experimentalanimals, primarily insects (Partridge & Mangel, 1999). Theresults of our study show weak evidence for late-age mor-tality deceleration in R. neglectus, in contrast to patternsreported for several Diptera, including the fruitfly Dro-sophila melanogaster Meigen (Curtsinger et al., 1992;Pletcher & Curtsinger, 1998), the medfly Ceratitis capitata(Wiedemann) (Carey et al., 1992, 1998), the mexfly Ana-strepha ludens (Lo¨w) (Carey et al., 2005), and the mosquitoAedes aegypti (L.) (Styer et al., 2007). Rhodnius neglectusage-specific mortality pattern seems to be more similar tothat reported for other kissing bugs such as: R. prolixus
Figure 3 Age-specific mortality rate l(x) of observed laboratorydata (as pool of five cohorts) of Rhodnius neglectus and the pre-
(Chaves et al., 2004a), R. robustus (Chaves et al., 2004b),
dictions of the DeMoivre, exponential, Gompertz, logistic, and
the milkweed bug, Oncopeltus fasciatus (Dallas) (Dingle,
Gavrilovs models, for the smoothed data applied to l(x).
In recent years, some striking lack of fit to the Gompertzmodel has been found in large cohorts of experimentalanimals, primarily insects (Partridge & Mangel, 1999). Theresults of our study show weak evidence for late-age mor-tality deceleration in R. neglectus, in contrast to patternsreported for several Diptera, including the fruitfly Dro-sophila melanogaster Meigen (Curtsinger et al., 1992;Pletcher & Curtsinger, 1998), the medfly Ceratitis capitata(Wiedemann) (Carey et al., 1992, 1998), the mexfly Ana-strepha ludens (Lo¨w) (Carey et al., 2005), and the mosquitoAedes aegypti (L.) (Styer et al., 2007). Rhodnius neglectusage-specific mortality pattern seems to be more similar tothat reported for other kissing bugs such as: R. prolixus
Figure 3 Age-specific mortality rate l(x) of observed laboratorydata (as pool of five cohorts) of Rhodnius neglectus and the pre-
(Chaves et al., 2004a), R. robustus (Chaves et al., 2004b),
dictions of the DeMoivre, exponential, Gompertz, logistic, and
the milkweed bug, Oncopeltus fasciatus (Dallas) (Dingle,
Gavrilovs models, for the smoothed data applied to l(x).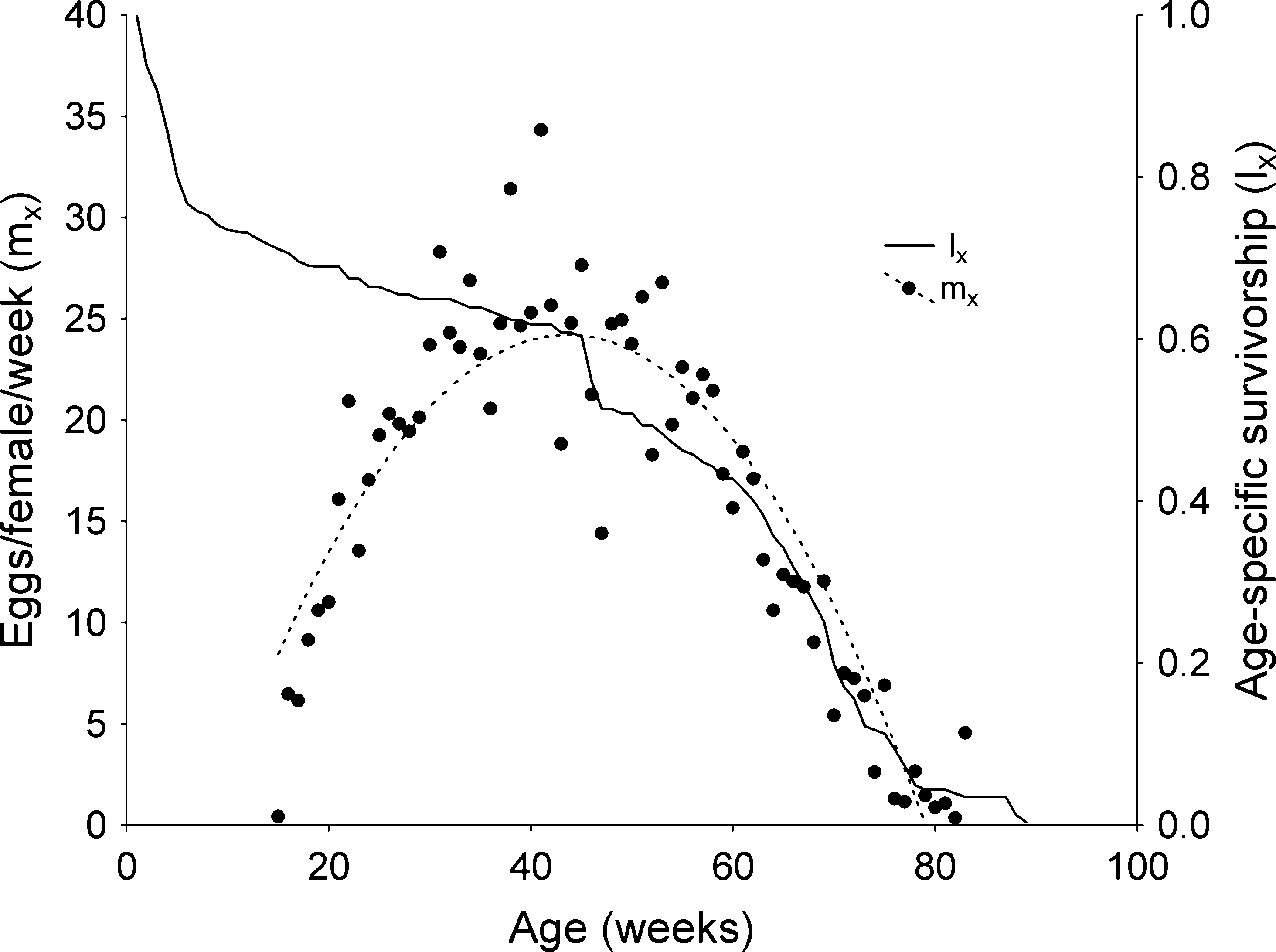 found in the beetle C. maculatus (Tatar et al., 1993) andthe kissing bug R. prolixus (Sulbaran & Chaves, 2006).
found in the beetle C. maculatus (Tatar et al., 1993) andthe kissing bug R. prolixus (Sulbaran & Chaves, 2006).